Species are kind of fuzzy. They become real over time, but it takes millions of years - James Mallet
It is surprising how little is formally proved about the origin of
species, given that was the title of Darwin’s seminal work and it is the major focus of evolution studies. There is of course a very large literature. The
subject is vitally important to establishing our own origins, and of necessity here only a brief primer of the most important results that might affect homo speciation are given.
The theory of speciation is simple enough. Different groups of the same
animal develop modifications through genetics that enable them to exploit
niches or differences in the environment, and they prosper, Eventually they
become so genetically separate from members of their species with other modifications that
they are unable to breed with them and they are then forever separate, building
their own variations.
We expect that speciation will happen very gradually as different characteristics are incorporated in separate populations. Along the way, we expect that hybridisation is initially possible and may occur repeatedly, but over time fewer and fewer individuals are able to mate successfully as the separate genetic characteristics become fixed.
Ring species and chronospecies
One of the major criticisms of evolution by creation
scientists has been the inability of geneticists to explain or demonstrate
speciation. Unfortunately no-one has ever seen a species emerge, it takes too
long, and no-one has really been able to examine a breeding population to see what
happened genetically as it approached speciation.
Clues to the answer come from so called “ring species”. These
are “chains of geographically adjacent
populations, in which each population interbreeds freely within itself but
somewhat less freely with those adjacent to it, and the populations at the ends
of the chain do not interbreed when they come in contact. By the most common
definition the populations at the ends of the chain have become separate
species."
Only four forms of life have been considered as candidates –
the herring or larus gull superspecies complex, the salamanders of the Central Valley in California, the song sparrow
of the Sierra Nevadas in California, and the Greenish Warbler of the Himalayas.
[1]
| Greenish warbler subspecies around the Himalayas |
In a ring species, crossbreeding is substantially
reduced between subspecies farther away along the ring. What seems likely is
that only a few individuals from distant populations can mate with each other,
as an intermediate step on the way to full speciation.
Richard Dawkins (2004) states that ring species "are
only showing us in the spatial dimension something that must always happen in
the time dimension”. The “ring over time” or chronospecies happens as the breeding group slowly
modifies its genetic structure to include various adaptations. We don’t know
exactly what mutations prevent viable offspring from forming but eventually
some members of the population will have these and they will not be able to
breed with the original population or with descendants that have headed down an
alternative path.
I think this probably happened with the Neanderthals when
sapiens came in contact with them - some but not all of them could hybridise with their very distant cousins - which explains the relatively small admixture in Eurasian humans.
Spatial mechanisms
Evolution 101 describes
four possible scenarios for the formation
of distinct species, as shown in the diagram.
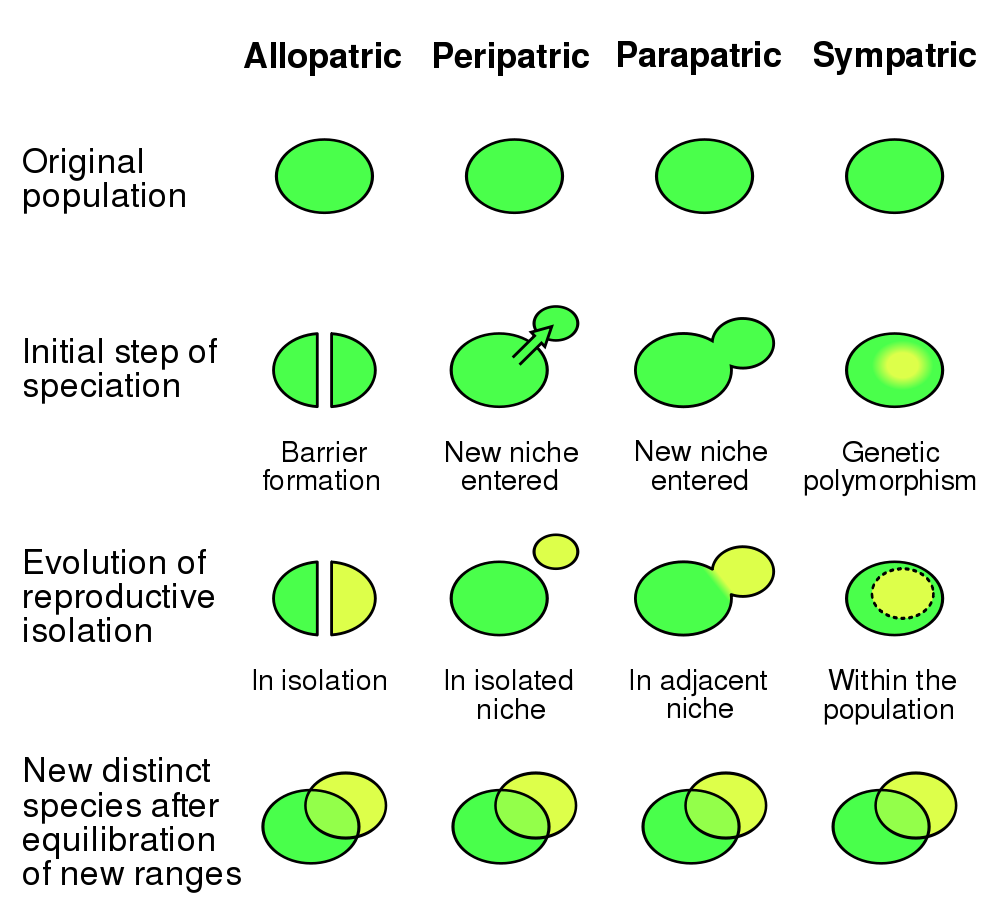
The first, "allopatric" scenario involving complete separation is straightforward - there is no mixing of genes and the populations develop independently. What is not quite clear is: how could two populations be so separated, unless they were on separate islands - and why did they not expand their range earlier if that were possible?
 |
| Allopatric speciation |
This is even more the case in the second "peripatric" scenario, which is the most favoured by evolutionists to be the usual path. The population enters a new niche and then is separated for some reason. It adapts and eventually forms a new species, then expands its territory to cross the original range. While this is a good scenario for the development of specific characteristics, it is hard to see where the separation could be maintained in hominims for the millions of years necessary for speciation. We will return to this in the human context.
The third "paripatric" scenario is essentially the ring species, or what happens with the different human "races" today. A gradient of different characteristics and (perhaps) differential breeding capacity forms across different locations. This definitely happens - but in practice there always seems to be sufficient gene leakage to prevent complete speciation.
The fourth "sympatric" possibility, in which connected populations diverge towards their intraspecies extremes to take advantage of local niches, remains speculative since no convincing cases have been observed. It requires both a very strong selection advantage at each extreme, and as well a mating preference among the differentiated individuals. This actually seems to be implied by "out of Africa" theory, since different hominim species would have to have both developed and subsequently lived in contact with each other.
Genetics and speciation
There are several well documented stages in species
separation, defined by the steps in production of viable offspring:
- sperm cannot fertilise eggs of other species
- eggs are fertilised but the embryos (particularly males) die
- viable offspring are produced but they are infertile
Species with different numbers of chromosomes generally can not pass step 1, because the chromosomes will not line up properly for fertilisation. This is particularly the case when males have more chromosomes than the females. However, in plants, there are
documented cases where hybrids of different species (eg radishes and cabbages)
can breed with each other but not the parent species. Sometimes this happens through a rare “bad” hybridisation of species with a different number of chromosomes that accidentally produces fertile offspring with some combined number of chromosomes. Similar phenomena have been observed in some insects and
fish.(Boxhom 1995)
“Behavioural” speciation has been frequently observed and can be easily induced in fruit fly. If separated groups develop different courtship behaviours they will not breed. This behavioural separation is sufficiently common that whether actual biological speciation has occurred can not be established “in the wild”, since the species refuse to breed.
The presence of different strains of parasite in some mosquitoes can prevent mating occurring – typically the egg and sperm nuclei fail to unite during fertilization. Treating the populations with antibiotics enables hybridization to occur.
Genetic quasi-speciation has occurred in some cases where the key gene for testicular development has moved on some individuals to another chromosome. Some of the hybrids will then be born without any copy of the gene and cannot produce males. This cannot happen if the gene is located on a non-recombinant Y chromosome, as in almost all mammals.
In mammals, breeding to form sterile mules or viable hybrids is quite common (see McCarthy for a long list). Horses, donkeys and zebras, each of which have different numbers of chromosomes, produce sterile hybrids. These have a common ancestor (MRCA) about ten million years ago. Lions, tigers and other Panthera species do the same. Several attempts to produce a chimpanzee-human hybrid (different number of chromosomes, MRCA about six million years ago) did not result in fertilization (Wikipedia, Mad Science).
The article Homo erectus sapiens stated our opinion that sapiens is not a species - because of fairly obvious hybridisation between sapiens and the other extant ssp of erectus with a common ancestor dating back a million years; and because the specific physical differences of sapiens do not appear to have obvious functional value - sapiens is simply erectus with a deformed head.
The arrival of erectus is a much clearer example of speciation, since they co-existed with australopithecines in Africa and possible elsewhere for such a very long time. Unfortunately we have no genetic evidence, and only a few bones on which to base hypotheses. Exactly how erectus was jump-started from pre-homo ancestors, and where it occurred, is anyone's guess.
These early erectus were functionally physically identical to us, apart from a somewhat smaller brain. They could therefore migrate long distances and, as far as we know, lived in a very wide variety of habitats. It is hard to imagine exactly how they could have engaged in peripatric speciation in an isolated location, unless they were trapped somewhere by rising sea levels and eventually escaped when an ice age opened paths.
This holds even more true for sapiens. It would take a long time to fix the distinctive brain and skull features in the absence of selective advantage. It is hard to think of anywhere in Africa that a race of people might have been separates from their kin for so long. Once again, they might have been isolated somewhere like the Indonesian archipelago and found their way off when boats were invented - or through a tsunami event.
Because so many species have different numbers of chromosomes and because this is known to limit fertility, chromosome splitting or combination may be a key requirement for speciation.
Speciation in the hominims
Although as we described in the introductory article, there are a plethora of species names for hominims that differ less from each other than we do among ourselves, there are in fact only two developments that might seriously qualify as speciation. One is the emergence of homo, especially home erectus, from the small-brained australopithecines before 1.6 million years ago. The second is the emergence of sapiens from erectus less than 200 000 years ago.The article Homo erectus sapiens stated our opinion that sapiens is not a species - because of fairly obvious hybridisation between sapiens and the other extant ssp of erectus with a common ancestor dating back a million years; and because the specific physical differences of sapiens do not appear to have obvious functional value - sapiens is simply erectus with a deformed head.
The arrival of erectus is a much clearer example of speciation, since they co-existed with australopithecines in Africa and possible elsewhere for such a very long time. Unfortunately we have no genetic evidence, and only a few bones on which to base hypotheses. Exactly how erectus was jump-started from pre-homo ancestors, and where it occurred, is anyone's guess.
These early erectus were functionally physically identical to us, apart from a somewhat smaller brain. They could therefore migrate long distances and, as far as we know, lived in a very wide variety of habitats. It is hard to imagine exactly how they could have engaged in peripatric speciation in an isolated location, unless they were trapped somewhere by rising sea levels and eventually escaped when an ice age opened paths.
This holds even more true for sapiens. It would take a long time to fix the distinctive brain and skull features in the absence of selective advantage. It is hard to think of anywhere in Africa that a race of people might have been separates from their kin for so long. Once again, they might have been isolated somewhere like the Indonesian archipelago and found their way off when boats were invented - or through a tsunami event.
Summary
Mammalian speciation has never been observed, but it seems to take place over an extended period where only some of the population are able to form hybrids with the main population from which they are diverging. Over time the proportion who would be able to mate successfully with the ancestral population goes down. At this stage, there is no evidence that erectus has ever been anything other than a single species; though it may be that different subspecies have hybridised with difficulty.Because so many species have different numbers of chromosomes and because this is known to limit fertility, chromosome splitting or combination may be a key requirement for speciation.
References
Dawkins, R. (2004).
The Ancestor's Tale, 2004:303
Mallet, J (2006). Subspecies, semispecies,
superspecies. Chapter In Encyclopaedia
of Biodiversity. (Elsevier).
[1] Unfortunately the first of these has turned out to be a
complex of different species and the second has been shown to occasionally
hybridise at the end of the chain,
No comments:
Post a Comment